Молекула АТФ — что это и какова её роль в организме. Пути синтеза атф в организме Регулирование РНР и связанных с ними ферментов, поддерживает баланс по отношению друг к другу
Любой организм может существовать до тех пор, пока происходит поступление питательных веществ из внешней среды и пока продукты его жизнедеятельности выделяются в эту среду. Внутри клетки происходит непрерывный очень сложный комплекс химических превращений, благодаря которым из питательных веществ образуются компоненты тела клетки. Совокупность процессов превращения материи в живом организме, сопровождающихся постоянным ее обновлением, и называется обменом веществ.
Часть общего обмена, которая состоит в поглощении, усвоении питательных веществ и создании за их счет структурных компонентов клетки, называется ассимиляцией - это конструктивный обмен. Вторую часть общего обмена составляют процессы диссимиляции, т.е. процессы разложения и окисления органических веществ, в результате которых клетка получает энергию, - это энергетический обмен. Конструктивный и энергетический обмен составляют единое целое.
В процессе конструктивного обмена клетка из довольно ограниченного числа низкомолекулярных соединений синтезирует биополимеры своего тела. Биосинтетические реакции протекают при участии разнообразных ферментов и требуют затрат энергии.
Живые организмы могут использовать только химически связанную энергию. Каждое вещество обладает определенным запасом потенциальной энергии. Главными материальными носителями ее являются химические связи, разрыв или преобразование которых приводит к освобождению энергии. Энергетический уровень одних связей имеет величину 8-10 кДж - эти связи называются нормальными. В других связях заключена значительно большая энергия - 25-40 кДж - это так называемые макроэргические связи. Почти все известные соединения, обладающие такими связями, имеют в своем составе атомы фосфора или серы, по месту которых в молекуле и локализованы эти связи. Одним из соединений, играющих важнейшую роль в жизнедеятельности клетки, является аденозинтрифосфорная кислота (АТФ).
Аденозинтрифосфорная кислота (АТФ) состоит из органического основания аденина (I), углевода рибозы (II) и трех остатков фосфорной кислоты (III). Соединение аденина и рибозы называется аденозином. Пирофосфатные группы имеют макроэргические связи, обозначенные значком ~. Разложение одной молекулы АТФ с участием воды сопровождается отщеплением одной молекулы фосфорной кислоты и выделением свободной энергии, которая равна 33-42 кДж/моль. Все реакции с участием АТФ регулируются ферментными системами.
Рис.1. Аденозинтрифосфорная кислота (АТФ)
Энергетический обмен в клетке. Синтез АТФ
Синтез АТФ происходит в мембранах митохондрий в процессе дыхания, поэтому все ферменты и кофакторы дыхательной цепи, все ферменты окислительного фосфорилирования локализованы в данных органеллах.
Синтез АТФ происходит таким образом, что два иона Н + отщепляются от АДФ и фосфата (Р) с правой стороны мембраны, компенсируя потерю двух Н + при восстановлении вещества В. Один из кислородных атомов фосфата переносится на другую сторону мембраны и, присоединив два иона Н + из левого отсека, образует Н 2 О. Остаток фосфорила присоединяется к АДФ, образуя АТФ.

Рис.2. Схема окисления и синтеза АТФ в митохондриальных мембранах
В клетках организмов изучено много биосинтетических реакций, использующих энергию, заключенную в АТФ, в ходе которых происходят процессы карбоксилирования и декарбоксилирования, синтеза амидных связей, образования макроэргических соединений, способных переносить энергию от АТФ к анаболическим реакциям синтеза веществ. Эти реакции играют важную роль в процессах обмена веществ растительных организмов.
С участием АТФ и других макроэргических нуклеозидполифосфатов (ГТФ, ЦТФ, УГФ) может происходить активирование молекул моносахаридов, аминокислот, азотистых оснований, ацилглицеринов путем синтеза активных промежуточных соединений, являющихся производными нуклеотидов. Так, например, в процессе синтеза крахмала с участием фермента АДФ-глюкозо-пирофосфорилазы образуется активированная форма глюкозы - аденозиндифосфатглюкоза, которая легко становится донором глюкозных остатков при формировании структуры молекул этого полисахарида.
Синтез АТФ происходит в клетках всех организмов в процессе фосфорилирования, т.е. присоединения неорганического фосфата к АДФ. Энергия для фосфорилирования АДФ образуется в ходе энергетического обмена. Энергетический обмен, или диссимиляция, представляет собой совокупность реакций расщепления органических веществ, сопровождающихся выделением энергии. В зависимости от среды обитания диссимиляция может протекать в два или три этапа.
У большинства живых организмов - аэробов, живущих в кислородной среде, - в ходе диссимиляции осуществляется три этапа: подготовительный, бескислородный и кислородный, в процессе которых органические вещества распадаются до неорганических соединений. У анаэробов, обитающих в среде, лишенной кислорода, или у аэробов при его недостатке диссимиляция протекает лишь в два первых этапа с образованием промежуточных органических соединений, еще богатых энергией.
Первый этап - подготовительный - заключается в ферментативном расщеплении сложных органических соединений на более простые (белков - на аминокислоты, жиров - на глицерин и жирные кислоты, полисахаридов - на моносахариды, нуклеиновых кислот - на нуклеотиды). Распад органических субстратов пищи осуществляется на разных уровнях желудочно-кишечного тракта многоклеточных организмов. Внутриклеточное расщепление органических веществ происходит под действием гидролитических ферментов лизосом. Высвобождающаяся при этом энергия рассеивается в виде теплоты, а образующиеся малые органические молекулы могут подвергнуться дальнейшему расщеплению или использоваться клеткой как «строительный материал» для синтеза собственных органических соединений.
Второй этап - неполное окисление (бескислородный) - осуществляется непосредственно в цитоплазме клетки, в присутствии кислорода не нуждается и заключается в дальнейшем расщеплении органических субстратов. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное расщепление глюкозы называют гликолизом.
Гликолиз - многоступенчатый ферментативный процесс превращения шестиуглеродной глюкозы в две трехуглеродные молекулы пировиноградной кислоты (пирувата, ПВК) С3Н4О3. В ходе реакций гликолиза выделяется большое количество энергии - 200 кДж/моль. Часть этой энергии (60%) рассеивается в виде теплоты, остальное (40%) используется на синтез АТФ.
В результате гликолиза одной молекулы глюкозы образуется по две молекулы ПВК, АТФ и воды, а также атомы водорода, которые запасаются клеткой в форме НАД Н, т.е. в составе специфического переносчика - никотинамидадениндинуклеотида. Дальнейшая судьба продуктов гликолиза - пирувата и водорода в форме НАД Н - может складываться по-разному. У дрожжей или в клетках растений при недостатке кислорода происходит спиртовое брожение - ПВК восстанавливается до этилового спирта:
В клетках животных, испытывающих временный недостаток кислорода, например в мышечных клетках человека при чрезмерной физической нагрузке, а также у некоторых бактерий происходит молочнокислое брожение, при котором пируват восстанавливается до молочной кислоты. При наличии в среде кислорода продукты гликолиза претерпевают дальнейшее расщепление до конечных продуктов.
Третий этап - полное окисление (дыхание) - протекает при обязательном участии кислорода. Аэробное дыхание представляет собой цепь реакций, контролируемых ферментами внутренней мембраны и матрикса митохондрии. Попав в митохондрию, ПВК взаимодействует с ферментами матрикса и образует: диоксид углерода, который выводится из клетки; атомы водорода, которые в составе переносчиков направляются к внутренней мембране; ацетилкофермент А (ацетил-КоА), который вовлекается в цикл трикарбоновых кислот (цикл Кребса). Цикл Кребса - это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуются две молекулы СО2, молекула АТФ и четыре пары атомов водорода, передаваемые на молекулы-переносчики - НАД и ФАД (флавинадениндинуклеотид). Суммарную реакцию гликолиза и цикла Кребса можно представить в следующем виде:
Итак, в результате бескислородного этапа диссимиляции и цикла Кребса молекула глюкозы расщепляется до неорганического диоксида углерода (СО2), а высвободившаяся при этом энергия частично расходуется на синтез АТФ, но в основном сберегается в нагруженных электронами переносчиках НАД Н2 и ФАД Н2. Белки-переносчики транспортируют атомы водорода к внутренней мембране митохондрий, где передают их по цепи встроенных в мембрану белков. Транспорт частиц по цепи переноса осуществляется таким образом, что протоны остаются на внешней стороне мембраны и накапливаются в межмембранном пространстве, превращая его в Н+-резервуар, а электроны передаются на внутреннюю поверхность внутренней митохондриальной мембраны, где соединяются в конечном итоге с кислородом.
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно, а снаружи - положительно (за счет Н), так что между ее поверхностями создается разность потенциалов. Известно, что во внутреннюю мембрану митохондрий встроены молекулы фермента АТФ-синтетазы, обладающие ионным каналом. Когда разность потенциалов на мембране достигает критического уровня (200 мВ), положительно заряженные частицы Н+ силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду.
Нормальное протекание метаболических реакций на молекулярном уровне обусловлено гармоничным сочетанием процессов катаболизма и анаболизма. При нарушении катаболических процессов прежде всего возникают энергетические трудности, нарушаются регенерация АТФ, а также поступление необходимых для биосинтетических процессов исходных субстратов анаболизма. В свою очередь, первичное или связанное с изменениями процессов катаболизма повреждение анаболических процессов ведет к нарушению воспроизведения функционально важных соединений - ферментов, гормонов и др.
Нарушение различных звеньев метаболических цепей неравнозначно по своим последствиям. Наиболее существенные, глубокие патологические изменения катаболизма происходят при повреждении системы биологического окисления при блокаде ферментов тканевого дыхания, гипоксии и др. или повреждении механизмов сопряжения тканевого дыхания и окислительного фосфорилирования (например, разобщение тканевого дыхания и окислительного фосфорилирования при тиреотоксикозе). В этих случаях клетки лишаются основного источника энергии, почти все окислительные реакции катаболизма блокируются или теряют способность аккумулировать освобождающуюся энергию в молекулах АТФ. При ингибировании реакций цикла трикарбоновых кислот выработка энергии в процессе катаболизма сокращается примерно на две трети.
В биологии АТФ - это источник энергии и основа жизни. АТФ - аденозинтрифосфат - участвует в процессах метаболизма и регулирует биохимические реакции в организме.
Что это?
Понять, что такое АТФ, поможет химия. Химическая формула молекулы АТФ - C10H16N5O13P3. Запомнить полное название несложно, если разбить его на составные части. Аденозинтрифосфат или аденозинтрифосфорная кислота - нуклеотид, состоящий из трёх частей:
- аденина - пуринового азотистого основания;
- рибозы - моносахарида, относящегося к пентозам;
- трёх остатков фосфорной кислоты.

Рис. 1. Строение молекулы АТФ.
Более подробная расшифровка АТФ представлена в таблице.
АТФ впервые обнаружили гарвардские биохимики Суббарао, Ломан, Фиске в 1929 году. В 1941 году немецкий биохимик Фриц Липман установил, что АТФ является источником энергии живого организма.
Образование энергии
Фосфатные группы соединены между собой высокоэнергетическими связями, которые легко разрушаются. При гидролизе (взаимодействии с водой) связи фосфатной группы распадаются, высвобождая большое количество энергии, а АТФ превращается в АДФ (аденозиндифосфорную кислоту).
Условно химическая реакция выглядит следующим образом:
ТОП-4 статьи которые читают вместе с этой
АТФ + Н2О → АДФ + Н3РО4 + энергия

Рис. 2. Гидролиз АТФ.
Часть высвободившейся энергии (около 40 кДж/моль) участвует в анаболизме (ассимиляции, пластическом обмене), часть - рассеивается в виде тепла для поддержания температуры тела. При дальнейшем гидролизе АДФ отщепляется ещё одна фосфатная группа с высвобождением энергии и образованием АМФ (аденозин-монофосфата). АМФ гидролизу не подвергается.
Синтез АТФ
АТФ располагается в цитоплазме, ядре, хлоропластах, в митохондриях. Синтез АТФ в животной клетке происходит в митохондриях, а в растительной - в митохондриях и хлоропластах.
АТФ образуется из АДФ и фосфата с затратой энергии. Такой процесс называется фосфорилированием:
АДФ + Н3РО4 + энергия → АТФ + Н2О

Рис. 3. Образование АТФ из АДФ.
В растительных клетках фосфорилирование происходит при фотосинтезе и называется фотофосфорилированием. У животных процесс протекает при дыхании и называется окислительным фосфорилированием.
В животных клетках синтез АТФ происходит в процессе катаболизма (диссимиляции, энергетического обмена) при расщеплении белков, жиров, углеводов.
Функции
Из определения АТФ понятно, что эта молекула способна давать энергию. Помимо энергетической аденозинтрифосфорная кислота выполняет другие функции:
- является материалом для синтеза нуклеиновых кислот;
- является частью ферментов и регулирует химические процессы, ускоряя или замедляя их протекание;
- является медиатором - передаёт сигнал синапсам (местам контакта двух клеточных мембран).
Что мы узнали?
Из урока биологии 10 класса узнали о строении и функциях АТФ - аденозинтрифосфорной кислоты. АТФ состоит из аденина, рибозы и трёх остатков фосфорной кислоты. При гидролизе фосфатные связи разрушаются, что высвобождает энергию, необходимую для жизнедеятельности организмов.
Тест по теме
Оценка доклада
Средняя оценка: 4.6 . Всего получено оценок: 522.
метаболиты гликолиза (1,3-дифосфоглицерат ,фосфоенолпируват ),
метаболиты цикла трикарбоновых кислот (сукцинил-КоА ) и
креатинфосфат .
Основным способом получения АТФ в клетке является окислительное фосфорилирование , протекающее в структурах внутренней мембраны митохондрий. При этом энергия атомов водорода молекул НАДН и ФАДН 2 , образованных в гликолизе, ЦТК, окислении жирных кислот,в ходе окислительно-восстановительных процессов преобразуется в энергию связей АТФ.
Однако также есть другой способ фосфорилирования АДФ до АТФ – субстратное фосфорилирование . Этот способ связан спередачей энергии макроэргической связи какого-либо вещества (субстрата) на АДФ. К таким веществам относятся:
Пируват окисляется до ацетил-КоА.
Пировиноградная кислота (ПК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. Ее судьба различна в зависимости от доступности кислорода в клетке. Ванаэробных условиях она восстанавливается домолочной кислоты . Ваэробных условиях пируват симпортом с ионами Н + , движущимися по протонному градиенту, проникает в митохондрии. Здесь происходит его превращение в ацетил-коэнзим А (ацетил-КоА ) с помощьюпируватдегидрогеназного мульферментного комплекса.
Пируватдегидрогеназный мульферментный комплекс

Суммарное уравнение окисления пировиноградной кислоты
Пируватдегидрогеназный мульферментный комплекс расположен в матриксе митохондрий эукариотов. Состоит у человека из96 субъединиц , организовавнных в три функциональных белка. Гигантское образование, имеет50 нм в диаметре, что впять раз!!! больше, чемрибосома .

Процесс проходит пять последовательных реакций, в которых принмает участие 5 коферментов:
Пируватдегидрогеназа (Е 1 , ПК-дегидрогеназа), коферментом служиттиаминдифосфат (ТДФ), катализирует 1-ю реакцию.
Дигидролипоил трансацетилаза (в русскоязычной литературе встречаются названия -дигидролипоат-ацетилтрансфераза илипоамид редуктаза трансацетилаза (Е 2), кофермент -липоевая кислота , катализирует 2-ю и 3-ю реакции.
Дигидролипоил дегидрогеназа (дигидролипоат-дегидрогеназа) (Е 3), кофермент –ФАД , катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими ферментами, в работе комплекса принимают участие коэнзим А иНАД .
Суть первых трех реакций сводится к декарбоксилированию пирувата (катализируется пируватдегидрогеназой, Е 1), окислению пирувата до ацетила и переносу ацетила на коэнзим А (катализируетсядигидролипоил трансацетилазой , Е 2).

Реакции синтеза ацетил-sКоА
Оставшиеся 2 реакции необходимы для окисления дигидролипоата обратно в липоат с образованием ФАДН 2 и восстановления НАДН (катализируютсядигидролипоил дегидрогеназой , Е 3).

Реакции образования надн Регуляция пируватдегидрогеназного комплекса
Регулируемым ферментом ПВК-дегидрогеназного комплекса является первый фермент – пируватдегидрогеназа (Е 1). Этому служат два вспомогательных фермента –киназа ифосфатаза, обеспечивая еефосфорилирования идефосфорилирования .
Киназа активируется при избытке конечного продукта биологического окисленияАТФ и продуктов ПВК-дегидрогеназного комплекса –НАДН иацетил-КоА . Активная киназа фосфорилирует пируватдегидрогеназу и инактивирует ее.
Фермент фосфатаза , активируясь ионамикальция или гормономинсулином , дефосфорилирует и активирует пируватдегидрогеназу.

Последующие этапы переваривания нерасщеплённого или частично расщеплённого крахмала, а также других углеводов пищи происходит в тонком кишечнике в разных его отделах под действием гадролитических ферментов - гликозидаз.
Панкреатическая α-амилаза
В двенадцатиперстной кишке рН среды желудочного содержимого нейтрализуется, так как секрет поджелудочной железы имеет рН 7,5-8,0 и содержит бикарбонаты (НСО 3 -). С секретом поджелудочной железы в кишечник поступает панкреатическая α-амилаза. Этот фермент гидролизует α-1,4-гликозидные связи в крахмале и декстринах.
Продукты переваривания крахмала на этом этапе - дисахарид мальтоза, содержащая 2 остатка глюкозы, связанные α-1,4-связью. Из тех остатков глюкозы, которые в молекуле крахмала находятся в местах разветвления и соединены α-1,6-гликозидной связью, образуется дисахарид изомальтоза. Кроме того, образуются олигосахариды, содержащие 3-8 остатков глюкозы, связанные α-1,4- и α-1,6-связями.
α-Амилаза поджелудочной железы, так же, как α-амилаза слюны, действует как эндогликозидаза. Панкреатическая α-амилаза не расщепляет α-1,6-гликозидные связи в крахмале. Этот фермент также не гидролизует (3-1,4-гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Целлюлоза, таким образом, проходит через кишечник неизменённой. Тем не менее непереваренная целлюлоза выполняет важную функцию балластного вещества, придавая пище дополнительный объём и положительно влияя на процесс переваривания. Кроме того, в толстом кишечнике целлюлоза может подвергаться действию бактериальных ферментов и частично расщепляться с образованием спиртов, органических кислот и СО 2 . Продукты бактериального расщепления целлюлозы важны как стимуляторы перистальтики кишечника.
Мальтоза, изомальтоза и триозосахариды, образующиеся в верхних отделах кишечника из крахмала, - промежуточные продукты. Дальнейшее их переваривание происходит под действием специфических ферментов в тонком кишечнике. Дисахариды пищи сахароза и лактоза также гидролизуются специфическими дисахаридазами в тонком кишечнике.
Особенность переваривания углеводов в тонком кишечнике заключается в том, что активность специфических олиго- и дисахаридаз в просвете кишечника низкая. Но ферменты активно действуют на поверхности эпителиальных клеток кишечника.
Тонкий кишечник изнутри имеет форму пальцеобразных выростов - ворсинок, покрытых эпителиальными клетками. Эпителиальные клетки, в свою очередь, покрыты микроворсинками, обращёнными в просвет кишечника. Эти клетки вместе с ворсинками образуют щёточную каёмку, благодаря которой увеличивается поверхность контакта гидролитических ферментов и их субстратов в содержимом кишечника. На 1 мм 2 поверхности тонкой кишки у человека приходится 80-140 млн ворсинок.
Ферменты, расщепляющие гликозидные связи в дисахаридах (дисахаридазы), образуют ферментативные комплексы, локализованные на наружной поверхности цитоплазматической мембраны энтероцитов.
Сахаразо-изомальтазный комплекс
Этот ферментативный комплекс состоит из двух полипептидных цепей и имеет доменное строение. Сахаразо-изомальтазный комплекс прикрепляется к мембране микроворсинок кишечника с помощью гидрофобного (трансмембранного) домена, образованного N-концевой частью полипептида. Каталитический центр выступает в просвет кишечника.
Сахаразо-изомальтазный комплекс. 1 - сахараза; 2 - изомальтаза;
3 - связывающий домен; 4 - трансмембранный домен; 5 - цитоплазматический домен.
Связь этого пищеварительного фермента с мембраной способствует эффективному поглощению продуктов гидролиза клеткой.
Сахаразо-изомальтазный комплекс гидролизует сахарозу и изомальтозу, расщепляя α-1,2- и α-1,6-гликозидные связи. Кроме того, оба ферментных домена имеют мальтазную и мальтотриазную активности, гидролизуя α-1,4-гликозидные связи в мальтозе и мальтотриозе (трисахарид, образующийся из крахмала). На долю сахаразо-изомальтазного комплекса приходится 80% от всей мальтазной активности кишечника. Но несмотря на присущую ему высокую мальтазную активность, этот ферментативный комплекс назван в соответствии с основной специфичностью. К тому же сахаразная субъединица - единственный фермент в кишечнике, гадролизующий сахарозу. Изомальтазная субъединица с большей скоростью гидролизует гликозидные связи в изомальтозе, чем в мальтозе и мальтотриозе.

Действие сахаразо-изомальтазного комплекса на мальтозу и мальтотриозу.

Действие сахаразо-изомальтазного комплекса на изомальтозу и олигосахарид.
В тощей кишке содержание сахаразо-изомальтазного ферментативного комплекса достаточно высокое, но оно снижается в проксимальной и дистальной частях кишечника.
Гликоамилазный комплекс
Этот ферментативный комплекс катализирует гидролиз α-1,4-связи между глюкозными остатками в олигосахаридах, действуя с восстанавливающего конца. По механизму действия этот фермент относят к экзогликозидазам. Комплекс расщепляет также связи в мальтозе, действуя как мальтаза. В гликоамилазный комплекс входят две разные каталитические субъединицы, имекдцие небольшие различия в субстратной специфичности. Гликоамилазная активность комплекса наибольшая в нижних отделах тонкого кишечника.
β-Гликозидазный комплекс (лактаза)
Лактаза расщепляет
β-1,4-гликозидные связи между галактозой
и глюкозой в лактозе.

Этот ферментативный комплекс по химической природе является гликопротеином. Лактоза, как и другие гликозидазные комплексы, связана с щёточной каемкой и распределена неравномерно по всему тонкому кишечнику. Активность лактазы колеблется в зависимости от возраста. Так, активность лактазы у плода особенно повышена в поздние сроки беременности и сохраняется на высоком уровне до 5-7-летнего возраста. Затем активность фермента снижается, составляя у взрослых 10% от уровня активности, характерного для детей.
Трегалаза - также гликозидазный комплекс, гидролизующий связи между мономерами в трегалозе - дисахариде, содержащемся в грибах. Трегалоза состоит из двух глюкозных остатков, связанных гликозидной связью между первыми аномерными атомами углерода.

Совместное действие
всех перечисленных ферментов завершает
переваривание пищевых олиго- и
полисахаридов с образованием моносахаридов,
основной из которых - глюкоза. Кроме
глюкозы, из углеводов пищи также
образуются фруктоза и галактоза, в
меньшем количестве - манноза, ксилоза,
арабиноза.

МЕХАНИЗМ ТРАНСМЕМБРАННОГО ПЕРЕНОСА ГЛЮКОЗЫ И ДРУГИХ МОНОСАХАРИДОВ В КЛЕТКИ
Моносахариды, образовавшиеся в результате переваривания, всасываются эпителиальными клетками кишечника с помощью специальных механизмов транспорта через мембраны этих клеток.
Всасывание моносахаридов в кишечнике
Транспорт моносахаридов в клетки слизистой оболочки кишечника может осуществляться разными способами: путём облегчённой диффузии по градиенту концентрации и активного транспорта по механизму симпорта за счет градиента концентрации ионов Na + . Na + поступает в клетку по градиенту концентрации, и одновременно глюкоза транспортируется против градиента концентрации (вторично-активный транспорт). Следовательно, чем больше градиент Na + , тем больше поступление глюкозы в энтероциты. Если концентрация Na + во внеклеточной жидкости уменьшается, транспорт глюкозы снижается. Градиент концентрации Na + , являющийся движущей силой активного симпорта, создаётся работой Nа + , К + -АТФ-азы, которая работает как насос, откачивая из клетки Na + в обмен на К + . В отличие от глюкозы, фруктоза транспортируется системой, не зависящей от градиента натрия.
Перенос в клетки слизистой оболочки кишечника по механизму вторично-активного транспорта характерен также для галактозы.
При разной концентрации глюкозы в просвете кишечника "работают" различные механизмы транспорта. Благодаря активному транспорту эпителиальные клетки кишечника могут поглощать глюкозу при её очень низкой концентрации в просвете кишечника. Если же концентрация глюкозы в просвете кишечника велика, то она может транспортироваться в клетку путём облегчённой диффузии. Таким же способом может всасываться и фруктоза. Следует отметить, что скорость всасывания глюкозы и галактозы гораздо выше, чем других моносахаридов.
После всасывания моносахариды (главным образом, глюкоза) покидают клетки слизистой оболочки кишечника через мембрану с помощью облегчённой диффузии в кровеносную систему.

 Организация наследственного материала у про-и эукариот
Организация наследственного материала у про-и эукариот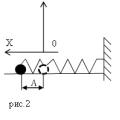 Пружинный маятник: амплитуда колебаний, период, формула Максимальная кинетическая энергия груза пружинного маятника
Пружинный маятник: амплитуда колебаний, период, формула Максимальная кинетическая энергия груза пружинного маятника Джек лондон биография на английском языке Джек Лондон биография на английском
Джек лондон биография на английском языке Джек Лондон биография на английском